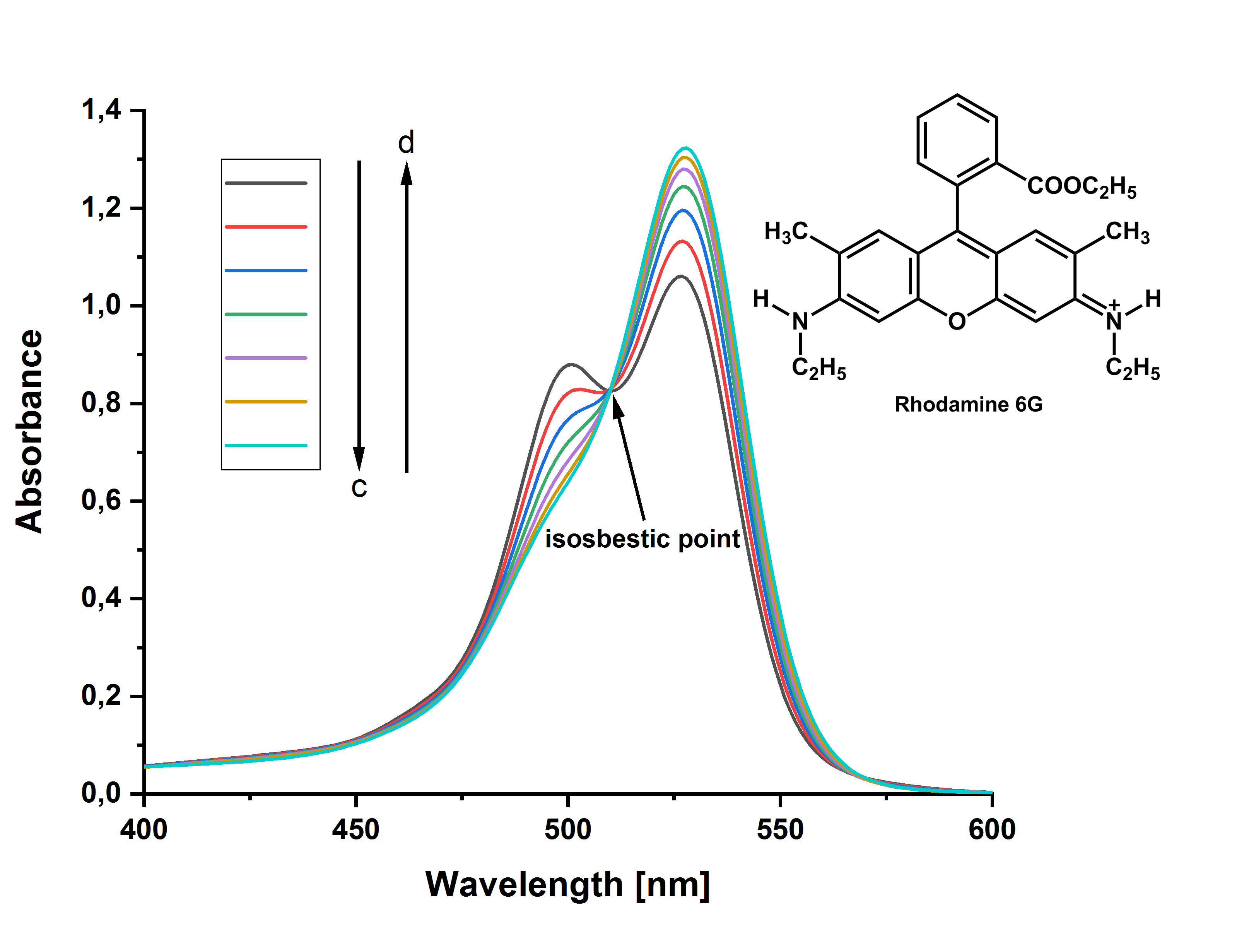
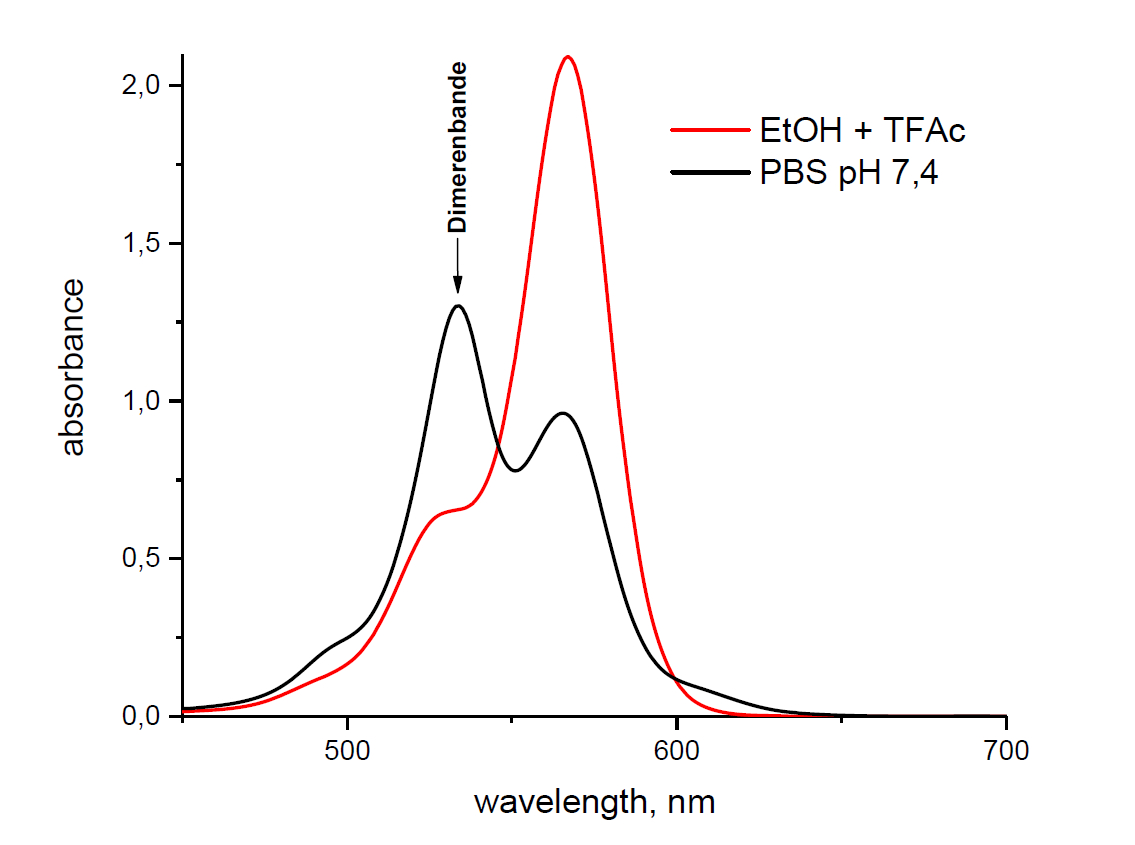
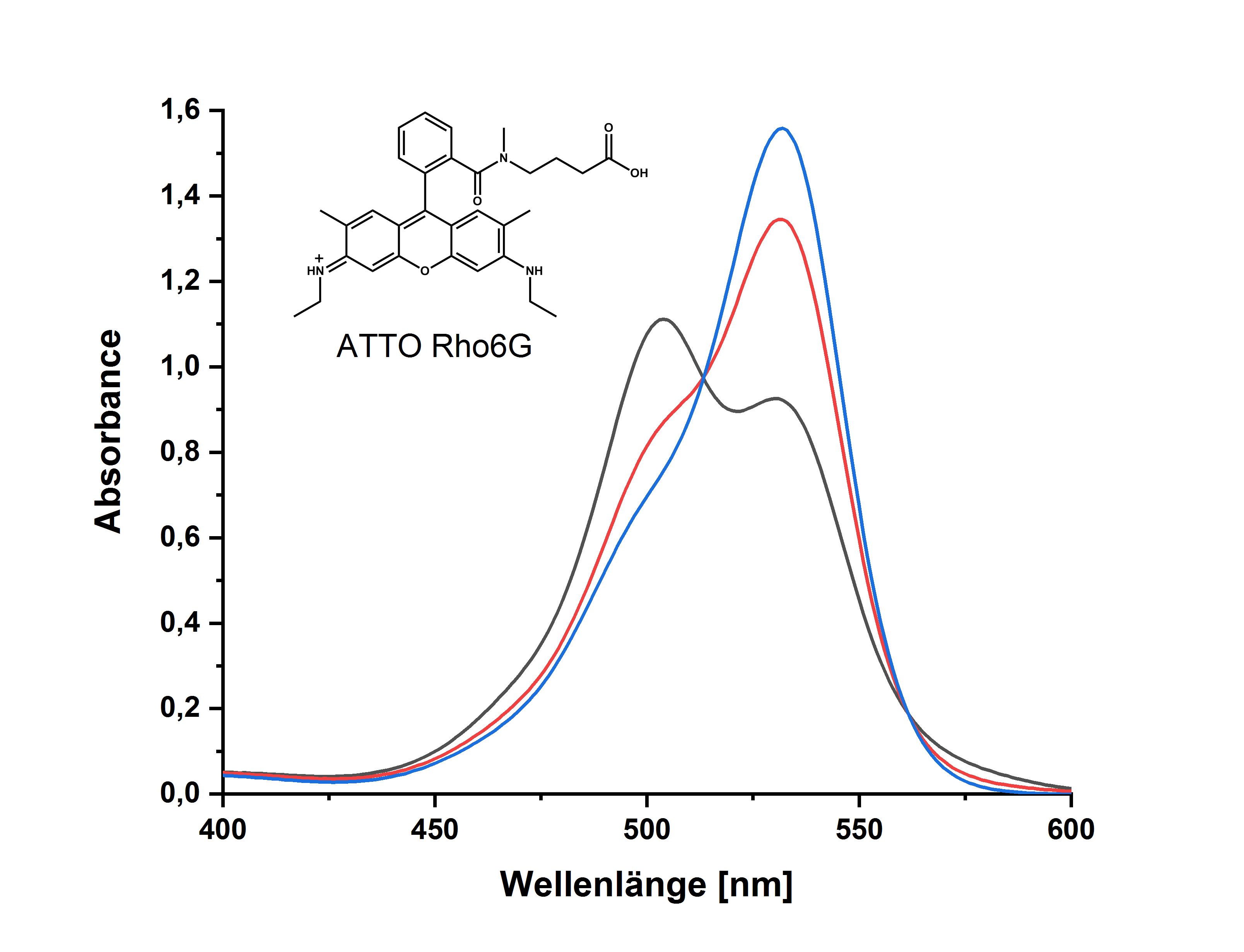
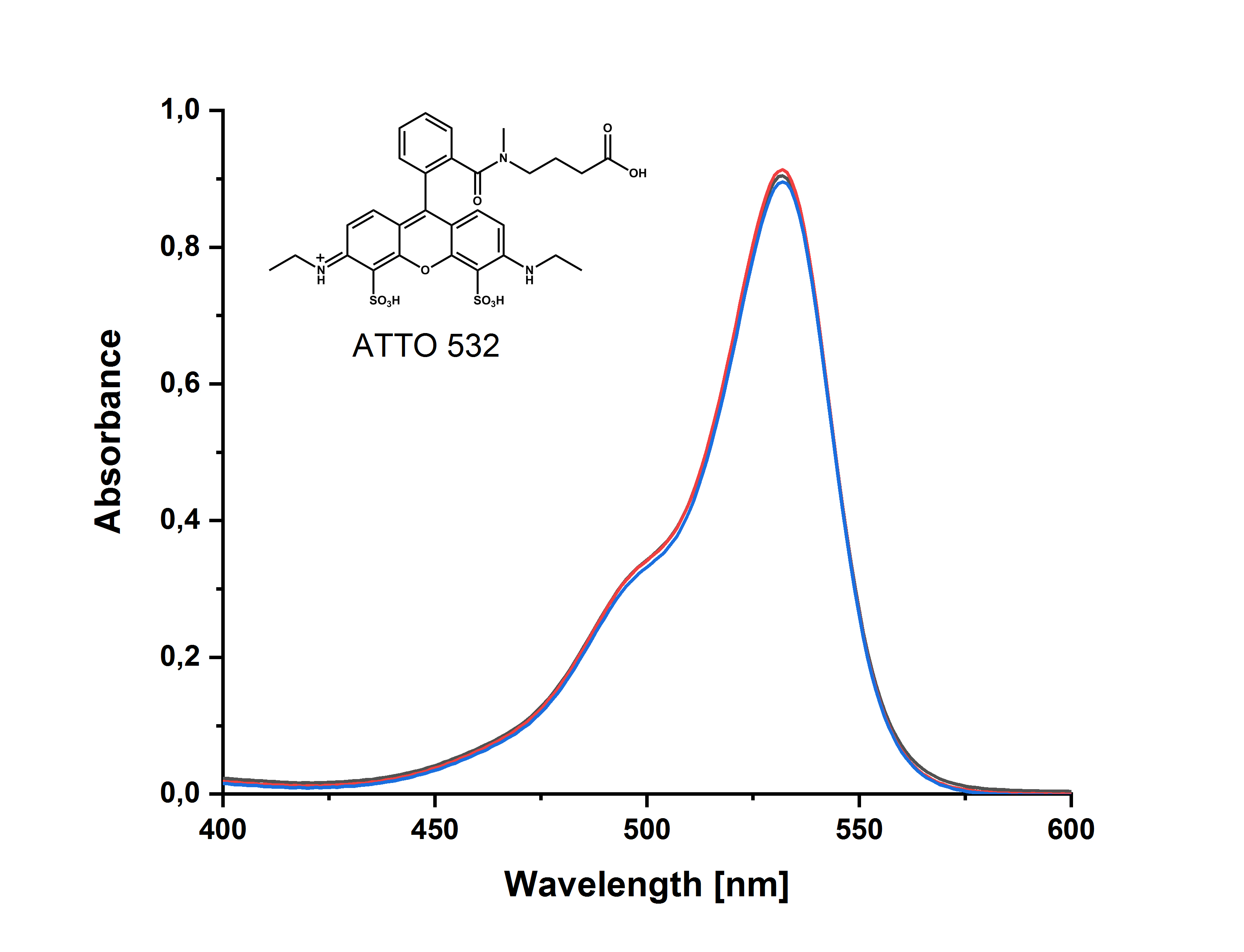
BOC Sciences小分子抑制剂介绍
抑制剂是能与蛋白质相互作用并降低靶蛋白生物活性的分子,包括酶抑制剂、转录因子抑制剂、离子通道阻滞剂、 等.小分子抑制剂的结构具有良好的空间分散性,其化学性质决定了良好的成药性和药代动力学性质。这些特性使小分子抑制剂在药物发现和开发过程中显示出很大的优势。因此,小分子抑制剂是一类潜在有用药物。我们的产品被纳入各种医学和制药研究领域,例如 神经系统疾病, 癌症, 代谢性疾病 等等。它们可以作用于流行的信号通路靶标,例如 PI3K/akt/mTOR, 细胞凋亡, 细胞周期/脱氧核糖核酸损伤, 贾克/统计 等等。
抑制剂的类型
-
不可逆抑制 – 抑制剂通常通过共价连接结合酶,这不可逆地使其失活。抑制剂可以破坏酶的结构和功能
-
可逆抑制-A可逆抑制剂通过非共价,更容易逆转的相互作用使酶失活。与不可逆抑制剂不同,可逆抑制剂可以与酶解离。 然后可以恢复酶的活性。可逆抑制可分为以下几类:
竞争性抑制 – 当与底物分子非常相似的分子与活性位点结合并阻止实际底物结合时,就会发生竞争性抑制。
非竞争性抑制 – 当抑制剂在活性位点以外的位点与酶结合时发生。此备用位置称为变构位点。结果,活性位点被改变,底物和酶不再像锁和钥匙一样结合在一起。因此,酶不能催化反应。由于抑制剂不与底物直接竞争,因此增加底物水平对降低抑制剂的作用没有影响。
非竞争性抑制 – 非竞争性抑制剂仅与酶 – 底物复合物结合,而不与游离酶结合。
拮抗剂:与受体结合时不产生生物反应,而是阻断或降低激动剂的作用。它可能是竞争性的,也可能是非竞争性的。
阻滞剂:也称为受体拮抗剂。它们是与受体结合并阻断激动剂作用的配体。
激动剂:一种能够结合和激活受体的药物,导致可能模仿天然物质的药理反应。它可以分为全部,部分或反向。
小分子抑制剂的应用
小分子抑制剂可以在自然界中找到,也可以在实验室中人工生产。天然存在的抑制剂对生命至关重要,并调节许多代谢过程。此外,自然产生的毒物通常是抑制剂,已经进化为对抗捕食者、猎物和竞争生物的有毒剂。 人工抑制剂常用作药物、杀虫剂和除草剂。
-
用作治疗疾病的药物
这是抑制剂最常见的用途,因为它们靶向人类酶并试图纠正病理状况。 大多数药物治疗各种慢性和危及生命的疾病,因为它们的特异性和可以抑制的酶的效力。抑制剂用于筛查推动抑制剂生长的各种疾病。
-
用作杀虫剂和除草剂
许多杀虫剂和除草剂都是酶抑制剂。从昆虫到人类,动物都含有一种叫做乙酰胆碱酯酶 (AChE) 的酶,这种酶通过将神经递质乙酰胆碱分解成乙酸盐和胆碱的机制,对神经细胞功能至关重要。许多 AChE 抑制剂用于医药和农业工业。它也被有机磷农药不可逆地抑制。
-
控制新陈代谢
抑制剂在代谢控制中发挥作用。细胞中的许多代谢途径受到控制酶活性的代谢物的抑制。这些代谢物通过底物抑制的变构调节来调节酶活性。
-
天然毒物质
植物和动物的进化必然会导致它们产生大量有毒物质,例如充当抑制剂的次级代谢物、肽和蛋白质。天然毒素通常是有机小分子,而且种类繁多,大多数代谢过程可能都有天然抑制剂。这些天然抑制剂不仅针对酶,而且还针对结构蛋白功能和受体通道。
-
分析传感器
这些传感器有助于监测各种环境因素。了解酶的抑制和再生机制对许多研究人员来说非常重要,尤其是在使用固定化酶时。
小分子抑制剂的优点
小分子抑制剂使用指南
作为治疗许多疾病的有前途的治疗分子,小分子抑制剂可以与其他治疗结合使用,例如RNA干扰(RNAi)和其他化学生物学探针。然而,确定与疾病相关的最特异性小分子抑制剂仍然很困难。我们提供了一些指南,以帮助您确定对其预期靶标具有高亲和力的优化抑制剂:
-
查看相关参考资料。作为参考,一些抑制剂可能具有特定的选择性和效力,作为药物或作为化学生物学研究的工具。
-
评估抑制剂是否在体外抑制靶标。合理的关系应该是体外生化测量的亲和力和有效细胞浓度之间的关系。它在文献中定期回顾或通过体外实验获得。
-
测量抑制剂的有效性。尽可能使用低浓度的抑制剂,因为浓度高于 10μM 时,抑制剂很有可能会抑制预期目标以外的其他物质。
-
观察是否有类似物抑制预期目标。有时,共享相同支架的相似抑制剂对细胞的效力与它们在生化测定中的效力相同。
-
在每个实验中使用阴性对照。生化测定中密切相关的类似物的失活。
-
使用阳性对照。为了证明抑制剂的选择性,有必要证明不同结构的抑制剂对细胞的效力与它们在生化测定中的效力相同。
-
救援实验。排除脱靶效应,挽救小分子诱导的表型是一种非常有用的方法。
其他小分子抑制剂选择指南
由于每个新的小分子都考虑用于您的研究,因此必须注意以下事项。
结构 应予以确认,其合成应可重现,以避免常见的毒性部分和泛测定干扰部分 (PAINS)。这也避免了化学反应性基团。
稳定性 应保存在相关媒体中。应注意任何pH敏感性。活动应保留在培养基中。分子不应表现出非特异性化学反应性,如氧化还原反应或膜不稳定。
溶解度 在水性介质中应足够。它还需要平衡溶解度和亲脂性。高可溶性和带电化合物可能显示出低细胞或组织通透性。疏水性化合物可能表现出高效率和渗透性,同时存在溶解度问题。使用盐形式的疏水分子应提高水溶性。
渗透性 通过被动手段或主动运输对于细胞测定中的活性是必需的。
IC50.半个最大抑制浓度(IC50)是衡量化合物抑制生物/生化功能的有效性的指标。在酶抑制过程中,IC50 表示在给定的实验条件下将酶促反应速率降低50%所需的抑制剂浓度。
K我 是描述抑制剂和酶之间结合亲和力的解离常数。双 IC50 和 Ki 值可用作抑制剂效力的定量指标。然而 Ki 是亲和力的内在量度,与酶浓度无关; 而 IC50 是描述特定抑制物质有效强度的操作参数,其取决于酶浓度。
体外和体内效力。体外效力通常单独使用,或与临床前体内数据结合使用从数据库和文献来源中提取药物体外效力参数(K d、K i、IC 50和 EC 50 )。
剂量依赖性活性通常是研究人员在先导药物原型中寻找答案的第一个标准。
构效关系 (SAR) 分析是一种旨在发现所研究化合物的化学结构(或结构相关特性)与生物活性(或目标特性)之间关系的方法。
可以设计阴性对照实验来证明抑制剂在用于抑制所需靶标的浓度下不会有效地改变任何脱靶蛋白的功能。
可以设计阳性对照实验来证明抑制剂在效果不明显的情况下按预期发挥作用。
作用机制是指药物产生药理作用的特定生化相互作用。了解药物的作用机制可能有助于提供有关药物安全性及其对身体影响的信息。它还可能有助于确定药物的正确剂量。

 另外,KPL新近推出8种DyLight荧光二抗,全面取代传统的FITC、Rhodamine、Texas Red、Cy3/5等荧光染料。他们在发光亮度、灵敏度上比传统的荧光抗体表现更好,背景更低;同时发光时间更持久,在不同pH及温度下具有更好的稳定性,更适合拍照。还有比FITC绿色荧光亮度更强、发光更持久、灵敏度更高、信噪比更低的绿色荧光标记吗?比Texas Red红色荧光亮度更强、发光更持久、灵敏度更高、信噪比更低的红色荧光抗体吗?
另外,KPL新近推出8种DyLight荧光二抗,全面取代传统的FITC、Rhodamine、Texas Red、Cy3/5等荧光染料。他们在发光亮度、灵敏度上比传统的荧光抗体表现更好,背景更低;同时发光时间更持久,在不同pH及温度下具有更好的稳定性,更适合拍照。还有比FITC绿色荧光亮度更强、发光更持久、灵敏度更高、信噪比更低的绿色荧光标记吗?比Texas Red红色荧光亮度更强、发光更持久、灵敏度更高、信噪比更低的红色荧光抗体吗?